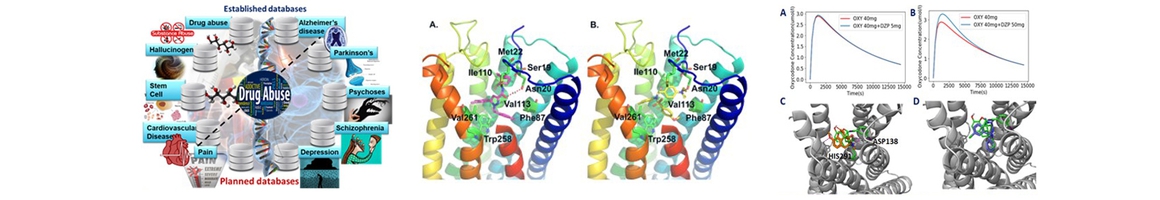
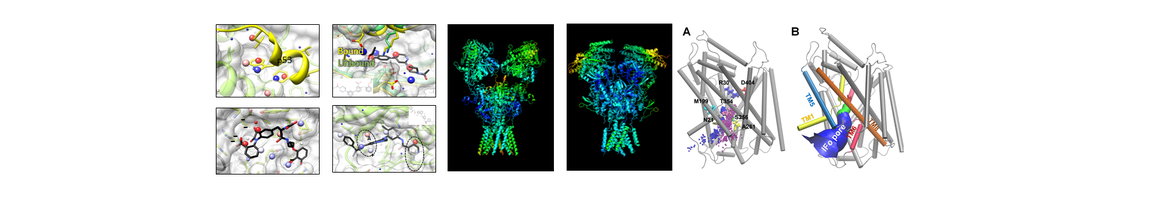
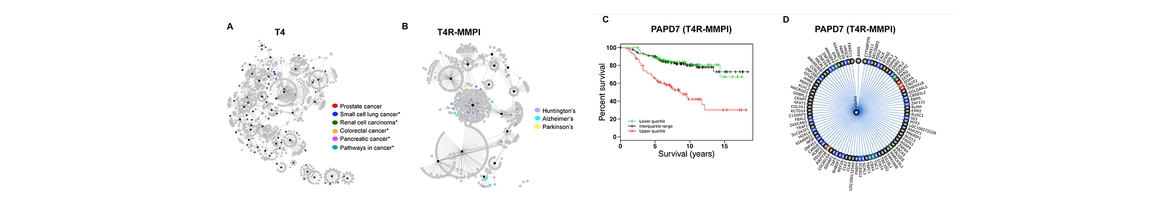


Role of Ubiquitin in Cardiovascular System
Li, Song
Kwon, Yong Tae
The N-end rule pathway is one ubiquitin proteolytic pathway that relates the in vivo half-life of a protein to the identity of its N-terminal residue. Conjugation of arginine (Arg) from Arg-tRNAArg to N-terminal aspartate (Asp), glutamate (Glu), or cysteine (Cys) is part of this proteolytic pathway in that it can lead to ubiquitination of the resulting Arg-conjugated proteins. We have previously identified the mammalian Ate1 gene encoding Arg-transferases responsible for all known protein arginylation activities and have shown that Ate1-/- embryos die owing to various cardiovascular defects including ventricular hypoplasia, ventricular septal defect, and late angiogenesis. These results suggest that Ate1-dependent proteolysis of unknown substrate(s) is a crucial regulatory mechanism for myocardial growth and blood vessel integrity/maturation. However, the exact nature of the cardiovascular defects and the underlying molecular mechanisms remain elusive. Genomewide functional proteomic approach led us to identify a set of cardiovascular regulators (Rgs4, Rgs5, and Rgs16) as substrates of Ate1-dependent arginylation that may underlie, at least partially, Ate1-dependent cardiovascular homeostasis. Notably, Rgs4 and Rgs5 are GTPase-activating proteins (GAP) that act as negative regulators of GPCR-coupled Ga subunits and have been implicated as important regulators of Gq/Gi-activated signaling for myocardial growth and vascular maturation/integrity, respectively. Biochemical analyses showed that degradation of these substrates depends on the Cys2 residue as a degradation determinant, which is exposed to the N-terminus through cleavage of N-terminal Met by Met aminopeptidases. In the presence of sufficient oxygen (O2) and nitric oxide (NO), N-terminal Cys2 appears to be oxidized to CysO2 to create a structural homolog of Asp, an arginylation-permissive residue. The N-terminal Arg residue of arginylated RGS proteins is subsequently bound by specific E3 ligases whose identities remain unclear. Using an affinity-based proteomic approach, we isolated a set of E3 family (named Ubr1 through Ubr7) and demonstrated that Ubr1, Ubr2, and Ubr4 are the major E3s specific for protein arginylation and that Ubr1-/-Ubr2-/- and Ubr4-/- embryos die of cardiovascular defects. Based on these results, we hypothesize that the functions of Rgs4, Rgs5, and Rgs16 are modulated through the MetAPs-O2/NO-Ate1-Ubr proteolytic cascade. In Aim 1, we will characterize the physiological function of Ate1-dependent arginylation in cardiovascular development and signaling using tissue-specific Ate1 knockout mice in combination with transgenic mice overexpressing Gq in the heart. In Aim 2, to understand the molecular principles underlying Ate1-dependent cardiovascular development, we will characterize arginylation-dependent turnover and cotranslational modifications of Cys2 of Rgs4 and Rgs5. In Aim 3, as part of our long-term efforts to characterize ubiquitin ligases specific of arginylated substrates, we will characterize cardiovascular development of mice lacking Ubr4, a newly identified recognition component downstream of protein arginylation. 1 PUBLIC HEALTH RELEVANCE: Cardiovascular processes involve many proteins whose functions can be turned on and off by posttranslational conjugation with compounds that are not used as a building block in protein (e.g., phosphate and GTP). We have previously shown that posttranslational conjugation of arginine, one of principal amino acids, to the protein N-terminus plays an important role in cardiac development and angiogenesis. In an independent study, we have demonstrated that Ate1-encoded R-transferases mediate proteasomal degradation of a set of cardiovascular regulators (Rgs4, Rgs5, and Rgs16) via the conjugation of arginine to the N-terminal Cys2 residue as a critical degradation determinant. Therefore, the arginylation of the Rgs proteins is a potential drug target to control Rgs-coupled G- protein signaling in the heart and blood vessels. Intriguingly, our more recent results suggest that Cys2- dependent proteasomal degradation of Rgs proteins requires not only arginylation but also oxidation, the latter being yet another unique modification as a licensing step prior to irreversible proteasomal degradation. The oxidation of the Cys2 residue of the Rgs proteins is in turn tightly controlled bybioavailability of molecules such as O2. The mammalian heart consumes O2 3-20 times more than the brain, and thus requires a constant supply of O2 for its function. O2 homeostasis is disrupted in heart disease, cancer, cerebrovascular disease, and chronic obstructive pulmonary disease, which represent the most common causes of mortality and accounts for two-thirds of all deaths in the U.S. Although O2 is a major determinant of cardiac gene expression and numerous cellular processes, extremely little is known about its role in the cardiovascular signaling and the mechanism by which the heart senses its concentration to modulate intracellular processes. Successful results from this study may provide a previously unknown mechanism sensing O2 and related molecules in response to a changing environment of circulating blood. Finally, our results suggest that the turnover of arginylated Rgs proteins is regulated by a set of E3 family (named Ubr1, Ubr2, and Ubr4) that recognize the N-terminal Arg through a structurally conserved zinc-finger domain, termed the Ubr box. The characterization of these Ubr E3 ligases will not only identify the physiological function of the arginylation-dependent proteolysis but also may reveal a new therapeutic target to control pathophysiological conditions in cardiovascular